![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

 Lichen biogeography
Lichen biogeography
Biogeography at different levels
You can look at lichen distributions at various taxonomic levels and it is useful to do so.
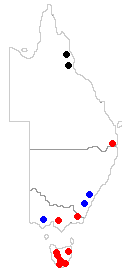 We'll start at the species level and consider four species. Metus conglomeratus has been found in parts of eastern Australia and New Zealand. The areas where it has been found in Australia are shown by the red dots on the map to the left. Myelorrhiza antrea and Neophyllis pachyphylla are known from limited areas of eastern Australia as you can see from that map (black dots for Myelorrhiza antrea and blue dots for Neophyllis pachyphylla) and Waynea hirsuta has been found only in the Tunkinksy Range near the southern end of Lake Baikal in central Siberia. The above evidence suggests that Metus conglomeratus evolved in Australasia and Neophyllis pachyphylla, being found only in widely separated areas of south-eastern Australia may have arisen there. When it comes to Myelorrhiza antrea it is harder to draw any conclusions since the species has been found only in such a limited area. The paper in which the species Myelorrhiza antrea was first described included the observation that "...it inhabits walls of caves where light can penetrate or in shaded rock crevices"
We'll start at the species level and consider four species. Metus conglomeratus has been found in parts of eastern Australia and New Zealand. The areas where it has been found in Australia are shown by the red dots on the map to the left. Myelorrhiza antrea and Neophyllis pachyphylla are known from limited areas of eastern Australia as you can see from that map (black dots for Myelorrhiza antrea and blue dots for Neophyllis pachyphylla) and Waynea hirsuta has been found only in the Tunkinksy Range near the southern end of Lake Baikal in central Siberia. The above evidence suggests that Metus conglomeratus evolved in Australasia and Neophyllis pachyphylla, being found only in widely separated areas of south-eastern Australia may have arisen there. When it comes to Myelorrhiza antrea it is harder to draw any conclusions since the species has been found only in such a limited area. The paper in which the species Myelorrhiza antrea was first described included the observation that "...it inhabits walls of caves where light can penetrate or in shaded rock crevices"![]() . Such places could easily be overlooked in a search for lichens so the species could well be more widespread, since much of tropical Queensland is still unexplored from a lichenological point of view. Similarly, based on the information given above, little can be said about Waynea hirsuta
. Such places could easily be overlooked in a search for lichens so the species could well be more widespread, since much of tropical Queensland is still unexplored from a lichenological point of view. Similarly, based on the information given above, little can be said about Waynea hirsuta![]() .
.
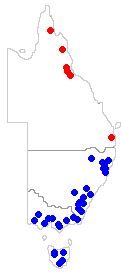 Let's go up a taxonomic level and look at what's known about the four genera. Within Myelorrhiza there are just two species (Myelorrhiza antrea and Myelorrhiza jenjiana) both so far found only in Queensland and the two species are similar in appearance. However, Myelorrhiza jenjiana grows on bark and has a wider known distribution, as shown by the red dots on the second map. As with Myelorrhiza there are just two species of Neophyllis and the second, Neophyllis melacarpa, is found in south-eastern Australia and is widespread in New Zealand. For interest the blue dots on the second map show the known eastern Australian distribution of Neophyllis melacarpa. Metus is a genus of three species and the remaining two (Metus efflorescens and Metus pileatus) are confined to southern South America. There are four more species of Waynea (Waynea adscendens, Waynea californica, Waynea cretica and Waynea stoechadiana). Waynea californica is known from western North America and the other three are found in the Mediterranean region
Let's go up a taxonomic level and look at what's known about the four genera. Within Myelorrhiza there are just two species (Myelorrhiza antrea and Myelorrhiza jenjiana) both so far found only in Queensland and the two species are similar in appearance. However, Myelorrhiza jenjiana grows on bark and has a wider known distribution, as shown by the red dots on the second map. As with Myelorrhiza there are just two species of Neophyllis and the second, Neophyllis melacarpa, is found in south-eastern Australia and is widespread in New Zealand. For interest the blue dots on the second map show the known eastern Australian distribution of Neophyllis melacarpa. Metus is a genus of three species and the remaining two (Metus efflorescens and Metus pileatus) are confined to southern South America. There are four more species of Waynea (Waynea adscendens, Waynea californica, Waynea cretica and Waynea stoechadiana). Waynea californica is known from western North America and the other three are found in the Mediterranean region![]() .
.
The known distribution of a particular species clearly gives essential information and, when used in conjunction with other facts (for example, habitat and climate data), may indicate where else that species might be found (or suggest ideas as to where it originated). Thus it would be reasonable to search for Myelorrhiza antrea in other caves or rock crevices of wet tropical Queensland. Taking a step up to the genus level helps put a species into a broader context. We see that both Myelorrhiza and Neophyllis are small genera but whereas the two species of the former have been found only in Australia, one species of the latter is widespread in New Zealand. This strongly suggests that the genus Neophyllis arose in Australasia, but where in Australasia is an open question, as is the evolutionary history of the two species. Thus far Myelorrhiza appears to be endemic to Australia but the occurrence of Myelorrhiza jenjiana further north makes one wonder if the genus could be found also in nearby Papua New Guinea, another region in need of further lichenological exploration. Of course the occurrence of that species in southern Queensland also suggests that the genus may be found along much of eastern Queensland.
|

Let's now turn to Metus. The current evidence suggests that Metus conglomeratus, confined to Australasia, arose there but the presence of two other species in South America raises the possibility that the genus had a Gondwanan origin. If so then there are questions such as where in Gondwana the genus arose and how did the currently known species evolve from the ancestral species. Of course, presence in both Australasia and South America does not by itself constitute proof of Gondwanan origin. Perhaps the generic ancestor species arose in one of Australasia or South America and dispersed, by wind say, to the other, after which time the isolated populations evolved independently to give rise to the species known today. The point is that, regardless of the true scenario, knowledge about the other species in the genus gives useful background information when it comes to investigating the biogeography of, say, Metus conglomeratus. Finally, it is worth noting that some lichenologists think that the differences between the Australasian Metus conglomeratus and the South American Metus pileatus warrant further investigation to see if there are good reasons for keeping the two species distinct. Should that not be the case we'd have just two Metus species, both found in South America and one in Australasia. Waynea, the last of the four genera, is predominantly Mediterranean but with two striking outliers. The evidence certainly suggests a Northern Hemisphere origin. One hypothesis to explain the current distribution of the genus is that the genus once was found in many more areas of the Northern Hemisphere but that changes in climate broke up a more continuous distribution, leaving separated populations to evolve into today's Mediterranean and outlier species.
The examples given above have involved small genera, each with just a few species. At the other extreme are large genera such as Xanthoparmelia, which has several hundred species. Though a cosmopolitan genus its greatest diversity is found in the southern hemisphere, in Australia and southern Africa, particularly in open habitats of the arid or semi-arid areas. It's clear that there are various groups of closely related species, where the species within a group are more closely related to each other than to any other species of Xanthoparmelia. The following diagram represents this sort of situation. Every dot indicates a species and the shorter the distance between two dots, the more closely related are the two species represented by those dots. You can see that the black dots indicate one group of closely related species, the blue dots another group and so on. The grey ellipse around the black and blue groups indicates that these two groups are closer to each other than either is to any other group of species and you could think of the grey ellipse as denoting a super-group. This grey super-group and the red group are closer to each other than either is to the brown group over on the left and the polygon marks out a still larger super-group.
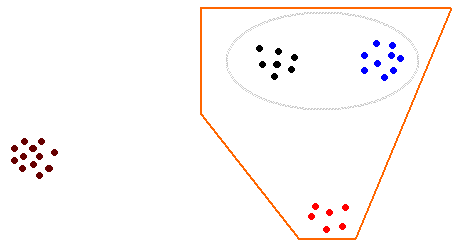
Depending on the question of interest a biogeographical analysis may focus on a species, a group, various super-groups or the whole genus. Thus it is reasonable to argue that all the species represented by the black dots are derived from a common ancestor (call it the black ancestor). Similarly we could posit a blue ancestor and, going further back in time an elliptical ancestor (that, via the black and blue ancestors, gave rise to today's black and blue species), a polygonal ancestor and so on. If it were possible to include a time scale, so that we knew roughly when the various ancestral species are likely to have existed, it may be possible to connect the evolution of the various groups of species to, say, various geological or climatic events.
The next step up the classification hierarchy is family. Just as a genus typically includes a number of related species, so a family typically includes a number of related genera. A very unusual species may be the only member in a genus and similarly an unusual genus may be the only member in a family. There's more about these topics under CLASSIFICATION. A family is essentially just another super-group and so introduces no new fundamental principles. Consequently, for the purposes of illustrating the variety of distribution patterns and the methods of biogeographical reasoning, species and genus will generally suffice in this introductory website.